Há quase uma década Casey Luskin escreveu réplica avassaladora contra um texto de senso comum escrito no Scienceblogs. A razão de depois de tanto tempo trazer o texto para o Portal vem do fato das críticas não “evoluírem” após a refutação. Repare na tentativa de falsear o que “não seria falseável”, a forte linguagem teleológica e o contentamento com homologias para inferir origem evolutiva.
Tradução por Enézio Eugênio de Almeida Filho.
O texto de Gabriel Cunha (UNIFESP), doutorando de Biologia Molecular, sobre os problemas da complexidade irredutível recomendado pelo Prof. Dr. Roberto G. S. Berlinck – Professor do Instituto de Química de São Carlos, da Universidade de São Paulo (USP), foi nocauteado no contexto de justificação teórica por um cruzado de esquerda do Design Inteligente (veja o texto dele aqui).
Autores de artigo do PNAS lançam mão de linguagem teleológica numa tentativa que falhou na explicação da evolução da complexidade irredutível.
Por Casey Luskin
Sumário: Um artigo publicado recentemente no Proceedings of the National Academy of Sciences (PNAS) tenciona explicar a evolução de uma máquina molecular relativamente pequena na mitocôndria que transporte proteínas pela membrana, desse modo supostamente refutando a complexidade irredutível. Frases e afirmações como “‘pré-adaptação’ para a bactéria antes de uma necessidade de importar proteína”, “as partes se acumulam até que estejam prontas para se encaixar”, “as maquinarias surgem antes de haver uma necessidade para elas” ou macromutações inteligentemente “engineered” [elaboradas] são inseparáveis desta última tentativa fracassada dos críticos do Design Inteligente (DI) de refutar o argumento de complexidade irredutível de Michael Behe. Como era de se esperar quando os evolucionistas são forçados a lançar mão de linguagem e mecanismos teleológicos orientados para objetivo, isto mostra que as explicações materialistas inerentemente cegas e não guiadas não são suficientes para produzirem sistemas de complexidade irredutível. Conforme discutido mais detalhadamente abaixo, esta última tentativa de refutar a complexidade irredutível mostra inconscientemente a necessidade de design inteligente. Um resumo dos problemas inclui:
- Nenhum proponente do DI jamais afirmou que esta máquina em particular seja de complexidade irredutível, então a refutação deles é do tipo “homem de palha” [espantalho]. - A maior parte do argumento é baseada na mera evidência de homologia de sequência sem nenhuma discussão detalhada de como evoluir realmente as proteínas bacteriais em máquinas eucarióticas de transporte mitocondriais, ou resolver outros problemas necessários para evoluir esta máquina biomolecular. - A maioria das questões sobre os passos evolucionários foi resolvida apelando-se para explicações aparentemente teleológicas que as proteínas foram magicamente “preadapted” [pré-adaptadas] para uso na máquina ou que as “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver necessidade para elas]. - Na sua única tentativa de usar uma proteína homóloga de uma bactéria no sistema eucariótico para testar a compatibilidade, os pesquisadores somente puderam fazer a função da proteína depois que eles “engineered” [elaboraram - palavra deles] inteligentemente as macromutações na proteína ao adicionarem extensas sequências de sinalização importantes bem como outras mudanças aparentemente necessárias. - Não houve evidência nenhuma de uma versão reduzida desta máquina de transporte (“TIM”) , por isso eles tiveram que tentar encontrar uma versão reduzida de uma máquina de transporte diferente, “TOM” para fornecer “proof of principle” [prova de princípio], mas não foram mencionados os homólogos procarióticos de TOM. - A versão supostamente mais simples de máquina de transporte TOM ainda requer múltiplas proteínas para funcionar, permitindo que ela ainda realize todas as funções essenciais, e o sistema tem pelo menos duas proteínas TOM – e não apenas uma como deu a entender o artigo do PNAS. - Foi afirmado que o sistema TOM supostamente mais simples foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas eram diferentes demais das proteínas TOM previamente conhecidas para serem detectadas. - Conforme a filogenia padrão, o precursor supostamente mais simples nem poderia representar uma forma ancestral porque ele descendeu de um ancestral que tinha os mecanismos típicos de transporte mitocondrial. Introdução
Artigos no SoftPedia e Wired estão trombeteando um artigo recente publicado no PNAS intitulado “The reducible complexity of a mitocondrial molecular machine” [A complexidade redutível da máquina molecular mitocondrial] afirmando que ele explica a evolução da complexidade irredutível. O parágrafo inicial do artigo do PNAS cita Michael Behe, declarando que “Os proponentes do Design Inteligente têm argumentado que estas máquinas sofisticadas são ‘irredutivelmente complexas’, servindo isto de prova que, a nível molecular, os princípios de evolução de Darwin não podem explicar a complexidade dos sistemas vivos”. É claro, os críticos do DI dizem constantemente que não deve existir nenhum debate científico sobre a questão da complexidade irredutível, mas não ligue pra isso.
Comecemos pelas afirmações do artigo da Softpedia sobre o flagelo bacteriano que os “os blocos básicos de construção deste ‘instrumento’ podem ser encontrados em outras partes na célula, mas realizando tarefas diferentes”. Por exemplo, todas as partes do flagelo não residem “em outras partes na célula” como um número de estruturas necessárias no flagelo sem homólogos fora do flagelo. E mesmo se todas as partes residissem em algum lugar na célula, só esperando para serem cooptadas para uso num flagelo bacteriano funcional, isto resolveria muito pouco para os evolucionistas. Angus Menuge explica por que estas estórias de coopção (também chamadas de “exapção”) são frágeis:
Para um flagelo funcional ser construído por exapção, as cinco condições seguintes deveriam ser todas atendidas: C1: Disponibilidade. Entre as partes disponíveis para o recrutamento na formação do flagelo, deveriam ser aquelas capazes de realizar as tarefas altamente especializadas de pá (tipo roda propulsora de navio), rotor, e motor, muito embora todos estes itens servem alguma outra função ou nenhuma função. C2: Sincronização. A disponibilidade destas partes teria que ser sincronizada de modo que em algum ponto, individualmente ou em combinação, todas elas estejam disponíveis ao mesmo tempo. C3: Localização. As partes selecionadas devem todas estarem disponibilizadas ao mesmo ‘sítio de construção’, talvez não simultaneamente, mas certamente na ocasião em que forem necessitadas. C4: Coordenação. As partes devem ser coordenadas somente no modo certo: mesmo se todas as partes de um flagelo estiverem disponíveis na ocasião certa, é claro que a maioria das maneiras de montá-las será não funcional ou irrelevante. C5: Compatibilidade de interface. As partes devem ser mutuamente compatíveis, isto é, ‘bem casadas’ e capazes de ‘interagirem’ apropriadamente: mesmo se uma pá (tipo roda propulsora de navio), rotor, e motor forem colocadas juntas na ordem certa, elas também precisam ter interface corretamente. (Angus Menuge, Agents Under Fire: Materialism and the Rationality of Science, p. 104-105 (Rowman & Littlefield, 2004).)
Os que se propõem explicar a evolução do flagelo quase sempre abordam somente a condição C1 e ignoram as condições C2-C5. Como será visto abaixo, esta mesma crítica se aplica com força igual aos argumentos presentes deste artigo do PNAS discutindo a origem dos mecanismos que transportam proteínas pela membrana mitocondrial.
A refutação de um homem de palha [espantalho]
As mitocondrias são um tipo de organela em células eucarióticas que produzem ATP, uma molécula que transporta eficazmente a energia utilizável para as reações celulares. Assim, as mitocondrias são frequentemente denominadas de “baterias” ou “casas de força” da célula. Elas são delimitadas por duas membranas externa e interna. Para se obter uma proteína fora da mitocondria para dentro da mitocondria, você deve passar a proteína pelas duas membranas através de vários tipos de mecanismos de transporte em cada membrana. Assim, a função destes mecanismos de transporte é pegar uma proteína e passá-la através de uma membrana. É uma função modesta, mas é uma função importante.
Basicamente, os autores do artigo do PNAS tentaram explicar como uma máquina molecular que transporta proteínas pela membrana interna da mitocondria, chamada TIM23, evoluiu. Nenhum proponente do DI jamais afirmou que este sistema particular é irredutivelmente complexo, tornando este artigo um ataque de homem de palha [espantalho]. Mas não ligue pra isso, certo?
A “Pré-adaptação” e as macromutações “elaboradas” inteligentemente resolvem as condições C2-C5?
Segundo os autores do artigo do PNAS, a evidência primária para a evolução do TIM23 é a mera semelhança entre seus componentes e outras proteínas que são encontradas em bactérias. Em um resumo-chave de seu argumento, que resolve somente a condição C1, os autores afirmam:
Nós propusemos que as máquinas “centrais” simples foram estabelecidas nos primeiros eucariotos pela utilização de proteínas bacteriais pré-existentes que previamente tinham fornecido funções distintas. Subsequentemente, e em um processo gradual de acordo com a evolução darwiniana, módulos adicionais teriam sido adicionados para as máquinas centrais para melhorar sua função. Esta proposição é apoiada por 3 descobertas: (i) que os componentes encontrados nas bactérias estão relacionadas em sequência aos componentes de máquinas de transporte de proteína mitocondrial, mas (ii) que estas proteínas bacteriais não são encontradas como parte das máquinas de transporte de proteína e (iii) que alguns organismos aparentemente “primitivos” encontrados hoje têm máquinas de transporte de proteína que funcionam somente com uma ou algumas partes componentes.
Como pode ser visto, o argumento deles se baseia principalmente em encontrar homólogos de proteínas de transporte mitocondrial em algum lugar na biologia. Então, eles explicaram como que os mecanismos de transporte mitocondrial evoluíram? Dificilmente. Isto explica apenas a condição C1. Mas, para oferecer verdadeiramente “a step-wise process in keeping with Darwinian evolution” [um processo gradual de acordo com a evolução darwiniana], eles devem explicar para as condições C2-C5 e explicar como que uma vantagem funcional foi obtida ao longo de cada pequena etapa daquela evolução.
Quanto a estes homólogos, a pesquisa publicada no PNAS descobriu que 2 proteínas em TIM23 — Tim44 e Tim14 — são similares a proteínas em bactérias, TimA e TimB. TimA e TimB têm uma função diferente nas bactérias (as bactérias não têm mitocondria), mas elas residem na membrana celular bacterial, algo do tipo como a Tim44 e Tim14 residem na membrana mitocondrial interna. Foi sugerido que, se TimA e TimB se combinassem espontaneamente com uma máquina transportadora de aminoácido LivH, seria possível evoluir um complexo transportador TIM23. Eis o resumo das etapas que eles propõem:
- A evolução de uma interação de ligação entre TimA e LivH que permita um ponto de encaixe para a proteína bacterial Hsp70, um motor transportador de proteína. - Depois a evolução das interações entre TimB e LivH. - Então, como é muito fácil fazer: "Com estas 3 proteínas bacteriais (TimA, TimB e LivH] cooperando como sub-unidades de uma máquina de transporte primitiva, será permitida a evolução gradual do complexo mitocondrial TIM."
O único aspecto real dos componentes explanadores requeridos de Menuge oferecido para as três proteínas é a condição C1. Eles afirmam que se as 3 partes estiverem disponíveis (etapa C1 de Menuge), então eles efetivamente resolveram o problema evolutivo. De modo algum. Quais são as probabilidades das partes se reunindo subitamente para formar um mecanismo de transporte mitocondrial funcional? Quais são as probabilidades de sítios produzindo subitamente ligação funcional entre todas estas proteínas? Estes são os principais problemas, mas eles tentaram pouco abordar estas questões. Na maior parte, eles apenas abordaram a condição C1 e declararam que o debate acabou. O argumento é algo como dizer que, se você colocar todas (ou neste caso, algumas das) partes de um computador numa caixa, e até sacudi-la um pouco, você obtém um computador. Mas as máquinas verdadeiras requerem muito mais do que apenas ter suas peças repousando numa caixa.
Há um modo de se determinar se as condições C2-C5 causam problema para a evolução destas proteínas. Eles poderiam substituir o DNA codificando Tim44 e Tim14 numa célula eucariótica de DNA com os genes codificando TimA e TimB, respectivamente, e verificar o que acontece.
Os autores não fizeram isto com TimA (eles apenas fizeram comparações sequenciais e estruturais) e, assim eles não deram realmente uma análise detalhada de como TimA pode evoluir em Tim44. “Mutações pontuais em um pequeno segmento exigido para a interação da proteína TimA com LivH forneceria um ponto de ancoragem para a Hsp70 bacterial, que é o homólogo direto do motor de transporte de proteína” se bem que eles não identificam quais seriam essas mutações pontuais, ou quantas seriam necessárias existir, ou quais outras mutações (tais como a inserção de sequências de sinalização apropriadas) seriam necessárias para evoluir um proteína do tipo TimA para funcionar como Tim44.
Para TimB evoluir em Tim14, a pesquisa deles é muito mais divertida. Os autores “engineered” (elaboraram – palavra deles) uma versão modificada de TimB, pela indução de mutações, não menos importante o fato de que TimB foi “elaborada para expressão em fermento adicionando-se uma sequência objetivando o terminal mitocondrial N e domínio de transmembrana”. Em outras palavras, eles solucionaram as condições C2, C3, e C4 – conseguindo a proteína expressada no lugar certo e no tempo certo através de algumas adições “engineered” [elaboradas] cuidadosa e inteligentemente completadas de sequências necessárias. Estes tipos de macromutações inteligentemente elaboradas não modelam o que a natureza poderia realizar razoavelmente.
Apesar disso, eles afirmam: “Embora a função da proteína bacterial em montagens simples, uma mutação relativamente pequena seria exigida para convertê-las em funcionalidade como uma máquina de transporte de proteína”. A expressão “mutação relativamente pequena” é um termo técnico? Adicionar espontaneamente as sequências-alvo adequadas, os domínios de transmembrana, e as sequências de sinalização, definitivamente não é uma “mutação relativamente pequena”. Se é que mostrou alguma coisa, esta pesquisa mostra é que a evolução funciona melhor quando é “engineered” [elaborada] inteligentemente.
Em vez de darem uma explicação cuidadosa das condições C2-C5, os autores do artigo do PNAS oferecem uma explicação que soa bem teleológica: “Juntas com o transportador de aminoácido LivH, estas partes componentes teriam fornecido a ‘pré-adaptação’ para as bactérias antes de uma necessidade de importação de proteínas”. Da mesma maneira, o artigo da revista Wired lida com este problema asseverando que estas partes que evoluíram, já estavam “prontas para se encaixarem” e subitamente realizaram totalmente uma nova função:
O processo pelo qual as partes se acumulam até que elas estejam prontas para se encaixarem é chamado de pré-adaptação. É uma forma de evolução neutra na qual o crescimento das partes não fornece nenhuma vantagem ou desvantagem imediats. A evolução neutra está fora das descrições de Charles Darwin. Mas, assim que as peças se reunem, a mutação e a seleção natural podem tomar conta do resto, finalmente resultando na agora complexa forma de TIM23].
A revista Wired até cita um dos cientistas que foi co-autor da pesquisa, dizendo, “Mas quando você pensa sobre isso de modo evolutivo neutralista, no qual estas maquinarias emergiram antes que houvesse necessidade para elas, então isto faz sentido”.
Faz realmente sentido que a refutação evolucionária para a complexidade irredutível seja simplesmente pura sorte, onde, absolutamente por nenhuma razão aparente as partes desta máquina só tiveram que ser “preadapted” [pré-adaptadas] e “prontas para se encaixar” e as “maquinarias que surgem antes que haja necessidade para elas”? Se isto é a resposta para Michael Behe, então parece que os argumentos de Behe ainda estão de pé.
Expressões como “ready to snap together” [prontas para se encaixar] ou “pre-adaptation” [pré-adaptação] soa altamente como dirigida a um objetivo, e um processo cego e não-dirigido como a evolução não deve ser evolução dirigida com objetivo. É claro que quando todas estas proteínas estiverem intactas e funcionando em conjunto, elas produzem uma função que dá uma vantagem. Mas, em primeiro lugar, de onde surgem subitamente estas partes, plenamente formadas, “ready to snap together” [prontas para se encaixar] para realizar esta nova função? Quando os evolucionistas invocam explicações do tipo “preadaptation” [pré-adaptação], Scott Gilbert parece estar muito certo em fazer a crítica que “[t]he modern synthesis is good at modelling the survival of the fittest, but not the arrival of the fittest” [a síntese moderna é boa em modelar a sobrevivência do mais apto, mas não a chegada do mais apto].
“Preadaptation” [pré-adpatação] ou “ready to snap together” [prontas para se encaixar] ou “machineries emerge before there’s a need for them” [maquinarias que surgem antes que haja necessidade para elas] não são explicações materialistas não-guiadas da evolução — gostem ou não, eles estão apelando inconscientemente para um processo dirigido com objetivo. E há somente um único processo dirigido com objetivo que eu conheço no pedaço: design inteligente.
Estas réplicas carregadas de teleologia me dão mais do que nunca confiança na potência do desafio da complexidade irredutível para a biologia evolutiva. São réplicas como estas que, em primeiro lugar, me fizeram cético do neodarwinismo e um proponente do Design Inteligente.
A máquina supostamente mais simples é a máquina errada e ainda exige muitas proteínas
Os autores do artigo do PNAS parecem reconhecer a improbabilidade destas partes estarem subitamente “ready to snap together” [prontas para se encaixar] e assim procuram simplificar o sistema, perguntando, “could a single component of the machine function in the absence of the others to provide even inefficient protein transport?” [poderia um só componente da função da máquina na ausência dos outros fornecer até mesmo o transporte ineficiente de proteína?] Aqui eles entram em dificuldades com respeito ao TIM23. Eles gostariam de encontrar um exemplo de apenas uma das proteínas TIM23 funcionando em algum lugar como um mecanismo de transporte pleno, mas eles não têm nenhuma evidência disso. TIM23 opera na membrana interior da mitocondria,mas existe um mecanismo de transporte similar na membrana exterior chamada TOM.
Através de uma pesquisa genômica, eles descobriram que em um caso, TOM parece ser composto de apenas 1 complexo de proteína, em vez dos costumeiros 3 ou 5 — ou como eles disseram (mais sobre isso abaixo). Isto dev ser considerado como “proof of principle” [prova do princípio] de que outras máquinas de transporte poderiam também funcionar com somente uma proteína:
Análise minuciosa da sequência do genoma de um grupo de organismos, a microsporidia, mostra que eles perderam os componentes deTom22, Tom5, Tom6,e Tom7 do seu complexo TOM central, e têm somente a sub-unidade de canal Tom40 (31).
Eles não relataram nenhum homólogo procariótico para proteínas no complexo TOM (alguém presumiria que eles teriam relatado se tivessem encontrado-os), assim, parece que eles não encontraram nenhum exemplo de mecanismos de transporte que sejam potencialmente redutíveis e tenham homólogos procarióticos. Apesar disso, os autores fizeram soar como se esta função possa ser realizada com apenas uma única proteína no complexo TOM, mas levante a referência 31 deles da citação acima, e você verá uma história muito diferente e muito mais interessante.
A citação dos autores do artigo do PNAS pra este ponto, a referência 31, é um artigo de janeiro de 2009 publicado no journal Eukaryotic Cell, mostrando que o sistema de transporte mitocondrial em certas microsporidia (um filo de parasitas unicelulares) é ainda muito complexo. (Nota: as microsporidia são muito diferentes de muitos outros eucarióticos, e têm uma organela tipo mitocondria chamada de mitosoma, mas nós seguiremos a terminologia do artigo do PNAS e apenas usar o termo “mitocondria” pelos propósitos desta réplica.) Olhe a figura 5 na página 6 do artigo (gratuito aqui). No lado esquerdo na parte A está o típico sistema de transporte mitocondrial conhecido da maioria dos eucarióticos; no lado direito na parte B está o sistema para o Encephalitozoon cuniculi, a espécie de microsporidia que, supostamente, tem um mecanismo de transporte mitocondrial reduzido.
Na figura 5A, repare que o típico sistema de transporte eucariótico mitocondrial tem máquinas de transporte redundantes (i.e. 2 tipos) membrana interior, mas somente um tipo na membrana exterior. Mas no E. cuniculi, há somente um mecanismo de transporte na membrana interior. Essencialmente, esta é a extensão da “complexidade redutível” que este artigo descobriu, conforme é visto na figura 1 abaixo:
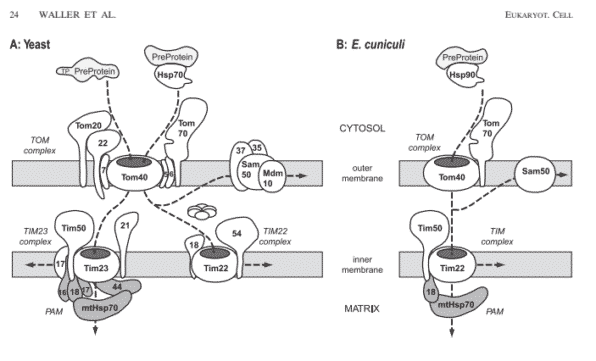
Tim23 e Tim22 têm funções e redundância que se sobrepõem para transportar proteínas através da membrana interior, como observa o artigo da Eukaryotic Cell, “As proteínas mitocondriais podem tomar uma das diversas rotas para os mitocôndrios através deste aparato” e os complexos aparecem em “forma especialista duplicada”. Parece não ser absolutamente necessário ter dois tipos de máquinas de transporte na membrana interior se uma pode realizar o trabalho. Portanto, é alguma surpresa encontrar um eucarioto mais simples que tem apenas um tipo de máquina de transporte na membrana interior? Isto não é uma descoberta surpreendente pois a maioria dos eucariotos superiores parecem ter sistemas redundantes na membrana interior.
Na verdade, com respeito a algumas das proteínas supostamente não encontradas no complexo TOM do E. cuniculi na membrana exterior, o artigo destaca que “substrate overlap between Tom20 and Tom70 occurs” [substrato se sobrepõe nos complexos Tom20 e Tom70 ocorre]. Outro estudo dos complexos TIM e TOM publicado no journal Current Biology declara do mesmo modo:
Os dois caminhos (das membranas exteriores) de targetting não são, estritamente, separados. Algumas pré-proteínas podem usar Tom70-Tom37 e Tom20-Tom22 e, mais importante, as pré-proteínas que são inicialmente reconhecidas pelo Tom70 são transferidas para Tom22 antes de sua inserção dentro do poro de importação formado por Tom40 (e possivelmente por Tom5]
Assim, há muita redundância no típico sistema de transporte eucariótico. Mas, olhando a figura 5b, no artigo, note que até no sistema “reduzido” no E. cuniculi, pelo menos 8 complexos de proteína (não uma proteína) ainda são necessários para realizarem todo o trabalho – inclusive os 4 componentes (não uma proteína) associados com a membrana exterior. E contra o artigo do PNAS, não existe apenas uma proteína no complexo TOM reduzido, mas duas.
Até onde eu sei nenhum proponente do Design Inteligente jamais afirmou que estes particulares mecanismos de transporte mitocondrial tenham complexidade irredutível. Mas, o que nós somos informados é que no E. cuniculi poderia representar um núcleo irredutível, como parece necessário para se ter uma máquina de transporte na membrana exterior e uma na membrana interior, cada uma com um número mínimo de partes para que este sistema funcione. O artigo do journal Eukaryotic Cell confirma este ponto ao afirmar:
Todavia, é possível que as nossas observações retratem uma maquinaria de importação que foi reduzida em resposta ao reducionismo dramático visto por toda a biologia microsporidiana. É notável que, da sub-série de maquinaria de importação que os HMMs identificaram, todas as principais funções essenciais estão representadas, embora somente uma vez em vez da forma especialista duplicada como é observado em fungos e animais (Fig. 5)”.
Eles estão dizendo que, embora nos eucariotos superiores exista redundância de função (“duplicate specialist form” [forma especialista duplicada]) para os sistemas de transporte, até em microsporidia, “all major essential functions are represented” [todas as principais funções essenciais estão representadas]. Assim, até com uma forma reduzida mais ainda complexa no E. cuniculi, ela é capaz de realizar todas as funções – uma grande admissão! Eles disseram mais que a configuração total mostra “evidence of a conserved system, albeit in relatively minimal form” [evidência de um sistema conservado, embora numa forma relativamente mínima]:
Nosso uso do HMM para pesquisar dados do E. cuniculi mostra que os candidatos para Tom70, Tom40, e Tim22 são identificados com grande confiança, e novos componentes adicionais do complexo TIM (Tim50 e Pam16) e o complexo SAM (Sam50) foram descobertos. Estes resultados oferecem mais evidência de um sistema de importação conservado, apesar de ser numa forma relativamente mínima (Fig. 5).
Realmente, quanto à proteína TOM70, eles afirmam, “Despite these losses, all other major structural features identified in the yeast Tom70 structure appear to have been conserved.” (emphasis added) [Apesar destas perdas, todas as principais características estruturais identificadas na estrutura Tom70 de fermento parecem ter sido conservadas (ênfase adicionada)]. Desse modo, não totalmente reduzido no sentido de que todos os principais componentes funcionais ainda estejam lá em alguma forma, e um punhado de proteínas são necessárias — na verdade, pelo menos 8 — para se obter uma proteína do exterior da mitocôndria para o interior da mitocôndria na microsporídia.
O artigo do PNAS constrói uma afirmação tipo homem de palha (espantalho) — que o sistema de transporte mitocondrial redundante em eucariotos superiores é de complexidade irredutível — e depois eles derrubam. O que nós observamos na microsporídia pode simplesmente representar um núcleo irredutível (vide aqui para uma explicação do conceito de “irreducible core” [núcleo irredutível]). Tente evoluir um sistema menos complexo do que este, e você pode não obter função nenhuma.
Na verdade, estes críticos do Design Inteligente podem ter deixado de fora do seu núcleo irredutível central um componente extremamente importante deste sistema — a sequência de sinal sobre as proteínas que serão transportadas. Assim como uma chave vira uma fechadura, um sinal sobre a proteína é tipicamente exigida para as proteínas ganharem acesso para dentro da mitocôndria através destes portais de transporte. Os autores também não consideraram a capacidade dos mecanismos darwinianos produzirem este componente fechadura-chave do sistema.
Assim, deve ser destacado que a referência 31 estuda o TOM, um sistema diferente do que aquele que o artigo do PNAS encontrou homólogos nas bactérias (TIM23). Olhando na referência 31, nós vemos o sistema de transporte mitocondrial mais simples conhecido é muito mais complexo do que o artigo do PNAS faz parecer.
Todas as proteínas TOM e TIM de E. cuniculi são atualmente detectáveis?
É importante salientar que o estudo no journal Eukaryotic Cell afirmou que o TOM no E. cuniculi foi reduzido somente após uma pesquisa genômica, que os pesquisadores admitem perderia proteínas que estavam realizando funções necessárias, mas que eram muito diferente das proteínas TOM previamente conhecidas como sendo detectadas. O E. cuniculi pode ter mais (ou até todas) as proteínas TOM padrão realizando todos os típicos papéis funcionais no sistema TOM típico, mas nós não saberíamos a partir desta pesquisa. Desta forma, o artigo do journal Eukaryotic Cell declara:
“A forma esqueletal da maquinaria de importação de proteína mitosomal identificada aqui pode retratar a dificuldade na identificação de homólogos de muitas das proteínas de importação dos fungos e animais, particularmente diversas das pequenas proteínas. Presentemente, poucos dados genômicos existem para o microsporidia com exceção do E. cuniculi, limitando a oportunidade de se procurar mais amplamente para os homólogos mitosomais neste grupo”. (ênfase adicionada)
Novamente, isto é uma grande admissão de que estas máquinas de transporte mitocondrial supostamente reduzidas possam muito bem ser mais complexas do que a pesquisa deles mostrou. Na verdade, o artigo observa que “many TOM and TIM proteins show weak conservation of primary sequence” [“muitas proteínas TOM e TIM mostram conservação fraca de sequência primária”]o que torna difícil identificá-las nas pesquisas genômicas. No caso de algumas das menores proteínas nos complexos TOM e TIM, o artigo debate se eles deveriam esperar encontrá-las:
“Embora algumas das menores proteínas (e.g., os pequenos Toms e os pequeníssimos Tims) possam ser difíceis de reconhecê-las por causa de sua relativa simplicidade destas sequências pequenas, a ausência de pares de muitas das proteínas de importação é conspícua considerando-se que elas são prontamente recuperadas de outros genomas de fungos e de animais”.
Em um caso eles escrevem que eles não têm certeza se a incapacidade de identificar certas proteínas nestes sistemas é devido ao fato de que elas não estão lá, ou se é porque eles são muito diferentes das proteínas equivalentes (que nós sequenciamos e estamos usando como uma base de pesquisa) nos eucariotos superiores:
“Não está claro se o fracasso em identificar mais complexos SAM de proteínas (Sam35, Sam37, and Mdm10) é devido à pobre conservação destas proteínas ou à capacidade de Sam50 de agir sozinho”].
Assim, mais proteínas equivalentes podem estar presentes no E. cuniculi do que foi relatado, mas elas podem não ter sido detectáveis na sua pesquisa genômica porque elas muito diferentes das proteínas de transporte atualmente conhecidas. Nós não sabemos com certeza qual é o caso porque estas pesquisas genômicas não investigam detalhadamente as atividades internas destes sistemas; eles apenas examinam as sequências de genes previamente conhecidas. Se as proteínas equivalentes estão no microsporidia, então não há razão para acreditar que ele tem um sistema de transporte mitocondrial reduzido.
O Encephalitozoon cuniculi não pode ser um precursor de eucariotos superiores
Finalmente, deve ser destacado que sob a filogenia de eucariotos prevalecente, é impossível que as máquinas de transporte de proteínas nos mitosomas do microsporidiano E. cuniculi representem algum estado antigo antes de os complexos múltiplos de TIM e TOM evoluírem em eucariotos superiores. Como o artigo no journal Eukaryotic Cell declara:
“Considerando-se que os microsporidia são mais proximamente relacionados com os fungos e, portanto, divergiram dentro da linhagem fungo-animal, o microsporidia ancestral muito provavelmente possuía as proteínas Tom e Tim comuns vistas hoje tanto nos fungos como nos animais”.
Deste modo, provavelmente os mecanismos de transporte reduzidos no E. cuniculi representam perda de função do estado normal mais complicado que nós encontramos nos eucariotos mais superiores:
“A especialização de complexos TIM no início da evolução eucariótica permitiu o tráfego mitocondrial de proteína. A presença de apenas um poro de membrana interior no E. cuniculi sugere que o microsporidia reverteu esta especialização devido ao tráfego reduzido de proteína e redução celular total”(ênfase adicionada).
Assim, este organismo não pode na verdade mostrar diretamente alguns precursores evolucionários para os mecanismos de transporte mitocondrial mais avançados que nós observamos hoje.
Conclusão
O artigo do Wired afirma que o sistema de transporte mitocondrial “seems to pose a cellular chicken-and-egg question: How could proteína transport evolve when it was necessary to survive in the first place?” [parece propor uma questão celular do tipo quem veio primeiro o ovo ou a galinha]. Contudo, como pode ser visto, as pesquisas estão mostrando que até nos sistemas “reduzidos” de transporte mitocondrial, “all major essential functions are represented” [todas as principais funções essenciais estão representadas ] e muitas proteínas são necessárias para realizar o trabalho. A principal diferença é a falta de sistemas redundantes e muito menos total de proteínas.
O artigo do Wired não resolveu esta “questão do tipo quem veio primeiro, o ovo ou a galinha” porque até o sistema tido como mais simples ainda é complexo, contendo máquinas múltiplas funcionais de transporte de múltiplas proteínas bem como de um número de proteínas adicionais requeridas para o processo de transporte.
O que é mais revelador é que esses evolucionistas foram forçados a lançar mão de linguagem explanatória dirigida a objetivos do tipo como “preadapted” [pré-adaptado], “parts accumulate until they’re ready to snap together” [as partes se acumulam até que estejam prontas para se encaixar]ou “machineries emerge before there’s a need for them” [as maquinarias surgem antes de haver uma necessidade para elas]. Nenhum defensor do DI jamais afirmou que esses sistemas particulares são irredutivelmente complexos, mas se esses argumentos dos evolucionistas dão alguma indicação, então mesmo aqui o Design Inteligente parece ter a vantagem, e os processos cegos e indiretos parecem insuficientes. Se a explicação deste artigo para a evolução desta máquina, em suas próprias palavras, “provides a blueprint for the evolution of cellular machinery in general” [fornece uma planta para a evolução da maquinaria celular em geral], estão está claro que as explicações evolutivas darwinianas para as origens da maquinaria celular são deficientes.
O artigo do PNAS afirma corretamente, “How these molecular machines evolved is a fundamental question” [Como que estas máquinas moleculares evoluíram é uma questão fundamental]. Alguém pensaria que uma questão fundamental dessas exigiria uma resposta rigorosa e detalhada. Infelizmente, os autores do artigo do PNAS tratam esta “questão fundamental” como se estivesse resolvida e a complexidade irredutível fosse refutada através de alguma investigação superficial e um punhado de explicações que parecem teleológicas. Eles querem ganhar o debate sem ter acontecido um.
Eles estão certos de que isto é uma “questão fundamental”, e a complexidade irredutível — nos casos onde os proponentes do DI têm argumentado realmente — ainda tem muito mérito.
Faça um comentário