Olá, Eskelsen! Eu tenho uma dúvida com relação a proteínas em um dos argumentos que usamos. A TDI tem um forte pilar que é mostrar que a evolução darwiniana não pode gerar novidade biológica (1), pois não pode gerar novas proteínas funcionais (novas funções). Isso porque a função de uma proteína está ligada ao seu sítio básico, e essas estruturas além de raríssimas, como a mostrada por Douglas Axe na ordem de uma em 10^77 (2), o sítio básico não funciona sem estar completo.
Até ai entendi perfeitamente.
Sei que uma proteína também pode sofrer adição de aminoácidos mas a função básica permanece a mesma. Porém o evolucionista não poderia dizer que essas adições gradativas em um sítio básico fizeram com que ao longo do tempo um novo sítio básico surgisse? (3)
Obs: Sei também que essa objeção só faz sentido se os sítios básicos de proteínas pudessem ser reduzidos a outros sítios com outra função. Porque se os sítios básicos são estruturas singulares, onde ao tirar aminoácidos não se chega a outros sítios mais simples, o evolucionista não teria para onde correr, pois não existe passo intermediário (4).
Se puder tirar essa dúvida vai ajudar muito.
David Balla
Segundo a definição de complexidade irredutível, é possível que várias enzimas tenham realmente algum grau de complexidade irredutível.
O processo evolutivo assegura que as estruturas biológicas possam ser formadas lenta e gradualmente por sucessivas mudanças que garantam alguma vantagem. Estruturas aparentemente simples como sítios ativos não estão inclusas na argumentação do design por complexidade irredutível, mas são considerados design em muitos casos por mera improbabilidade.
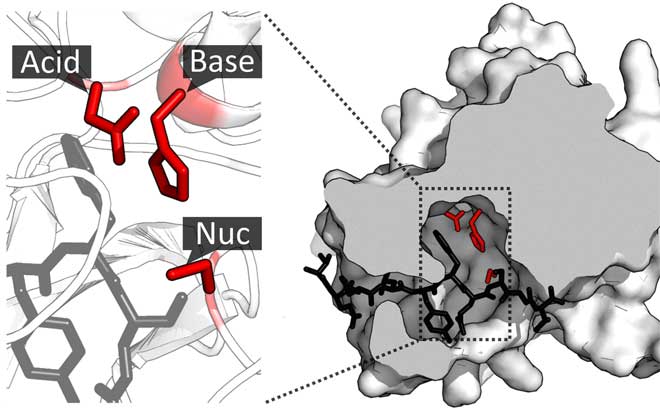
Os sítios ativos são regiões nas enzimas que favorecem reações químicas. Nem toda proteína é uma enzima, mas algumas possuem o sítio ativo como uma ferramenta para trabalhar sobre outras moléculas (substratos). E esse sítio ativo permite essa atividade enzimática, que é promover alguma reação química.
Sítios Ativos: quando proteínas são enzimas
Os organismos precisam manter suas estruturas biológicas e as enzimas trabalham nisso sem cessar. Elas compõe o que chamamos “metabolismo”. Elas tecem desde as unhas até os fios de cabelo, a pele, os tecidos em geral, fazem reparos; participam ativamente dos sentidos: transformam estímulos em sinais para que se possa ver, sentir sabores, ouvir etc. e etc. Praticamente para cada necessidade biológica há no mínimo uma proteína relacionada (quando não milhares).
O sítio ativo possui ainda internamente um sítio de ligação, para o encaixe, e o sítio catalítico, onde a atividade enzimática ocorre:

Como você pode ver, o sítio catalítico depende da conformação do sítio de ligação e este da conformação do sítio ativo, todos eles ainda dependem da estrutura tridimensional da proteína.
Cofator: a ferramenta para as atividades
Ainda há outro elemento crucial nas enzimas, o cofator. Substâncias inorgânicas necessárias para atividade enzimática. Íons de metais como cálcio, zinco, potássio, sódio, manganês, magnésio, cobre e ferro são ajustados no sítio catalítico para acelerar as reações. Algumas reações levariam até milhões de anos para acontecer, outras poderiam mesmo tomar bilhões de anos. As enzimas fazem essa reação acontecer em milissegundos.
Na perspectiva evolutiva, fenômenos como a promiscuidade enzimática combinada com duplicação de genes tentam explicar a origem das enzimas. A promiscuidade enzimática é a capacidade de uma enzima catalisar outras reações além de sua reação principal. Creio que o caso emblemático da nylonase ilustre bem isso, mas no fim é um exemplo modesto que não dá o suporte a narrativa evolutiva para a diversidade biológica.
Respondendo aos pontos
1. Sobre “a evolução darwiniana não pode gerar novidade biológica”. Na verdade, não é uma impossibilidade absoluta, mas uma grande improbabilidade que reduz totalmente a plausibilidade. É algo desfavorecido que inviabiliza toda a história evolutiva proposta. A evolução darwiniana é muito pobre em gerar novidade biológica de forma que as perdas superam qualquer ganho. A tendência é o declínio dos sistemas biológicos, o que refuta as pretensões darwinianas. Behe fez um vasto trabalho para averiguar até onde o darwinismo pode ir e conseguiu alguns raros exemplos limite que deram os contornos a sua hipótese:
Um exemplo de um verdadeiro ganho de FCT por duplicação citado em minha publicação foi o trabalho de Olsthoorn e van Duin (1996), onde a duplicação de 14 nucleotídeos levou à formação de novos elementos codificados funcionais (não simplesmente repetindo elementos preexistentes), por isso não é simplesmente uma mutação de modificação-de-função.
Michael Behe. Óculos com lentes cor-de-rosa: Lenski, citrato e a BioLogos.
2. A raridade de uma em 10^77 de Axe é sobre a dobra da estrutura tridimensional. O sítio ativo ainda será um ínfima fração dentro desse número, porque o sítio depende criticamente da estrutura tridimensional. Esse número é ainda desconhecido.
3. Não sei dizer ao certo quais casos um sítio ativo pode ser tornar outro através de seleção, mas sabemos que isso não elimina de fato a complexidade irredutível da estrutura. Um caso de refutação seria uma proteína sem sítio ativo que desenvolvesse essa estrutura, e isso se aplicaria para o caso específico a princípio sem ser generalizado.
4. Essa redução para estruturas mais simples é um problema evolutivo grave. Eles realmente não conseguem nada persuasivo. Afirmar que complexidade irredutível não existe e sofrer frustrações desde a década de 1970 tentando reduzir essas estruturas deve ser algo singular na história da ciência.
Ilhas Funcionais
Algumas funções estão próximas no espaço de possibilidades, a minoria. Então a mudança de uma função para outra, que não refuta a complexidade irredutível, não é algo dominante. Pelo contrário, as funcionalidades estão longe uma das outras, em “ilhas funcionais”. Isso significa que a passagem gradual de uma configuração para outra é implausível.
Texto Relacionado:
…
Faça um comentário